- research article
- https://doi.org/10.12345/biog.2026.0007
-
- VOL 3
- 2026
-
- Received:
- Accepted:
- Published:
Plain Language Summary
Island plants are not distributed randomly. Some islands preserve old climatic refuges where lineages survived environmental change for long periods, and those places often contain unique genetic variation and unusual combinations of traits. In this study, we show that islands with long-term climatic stability matter for conservation even when they do not contain the highest number of species today. That means biodiversity planning should consider deep history and ecological function, not just species counts.
Abstract
Oceanic archipelagos concentrate endemic lineages, steep environmental gradients, and high extinction risk, making them ideal systems for testing how climatic stability, dispersal limitation, and lineage history jointly shape biodiversity patterns. Yet integrative studies linking phylogeographic structure to functional turnover across entire island networks remain uncommon. We evaluated whether climatically stable upland refugia predict contemporary endemic richness, phylogeographic partitioning, and functional beta diversity across the Macaronesian archipelagos, with particular attention to endemic shrub assemblages spanning the Azores, Madeira, Selvagens, Canary Islands, and Cabo Verde. We assembled 412 georeferenced occurrence records for 37 endemic shrub taxa, generated species distribution models under contemporary and late-Holocene climate layers, summarized trait space using leaf area, seed mass, wood density, and maximum height, and reconstructed chloroplast haplotype networks for 18 focal taxa. We then linked local assemblage composition to island area, topographic complexity, refugial persistence, and inter-island isolation using generalized linear mixed models, variation partitioning, and distance-based redundancy analyses. Refugial persistence was the strongest predictor of endemic richness and unique haplotype occurrence, exceeding the explanatory power of island area alone. Functional turnover increased sharply along aridity and elevational contrasts, whereas taxonomic turnover saturated with geographic distance. Several islands retained deep phylogeographic splits despite modest present-day richness, indicating a decoupling between lineage history and contemporary species counts. Conservation prioritization based only on richness overlooked islands harboring disproportionately high evolutionary and functional uniqueness. Macaronesian endemic shrub assemblages reflect the combined influence of climatic microrefugia, historical isolation, and trait-mediated environmental filtering. Biogeographic prioritization should therefore integrate taxonomic, functional, and phylogeographic information rather than relying on richness metrics alone.
Introduction
Biogeography seeks to explain why organisms occur where they do, and why those patterns vary across space, time, and lineage history. Oceanic archipelagos have been central to that project since classic work on island assembly and turnover (MacArthur and Wilson, 1967). More recent developments in macroecology, phylogeography, and conservation science have broadened this agenda by asking how climatic instability, trait filtering, and population fragmentation interact across nested spatial scales (Whittaker et al., 2017); (Mittelbach and Schemske, 2015).
The Macaronesian archipelagos provide an especially informative test bed because they combine volcanic island ontogeny, strong gradients in aridity and elevation, repeated long-distance colonization, and uneven histories of land-use pressure. Previous work has shown that endemic plant diversity in these systems is shaped by both isolation and environmental heterogeneity, but the relative contribution of long-term climatic refugia remains unresolved (Fernández-Palacios et al., 2011). Narrative evidence from lineage-level studies suggests that persistent cloud-forest and montane habitats may preserve deeply divergent haplotypes, even where contemporary richness is modest (Emerson, 2002).
That problem matters beyond regional natural history. Conservation biogeography increasingly emphasizes that richness alone can misidentify priority areas when evolutionary history and functional distinctiveness are spatially decoupled (Pollock et al., 2017; Tucker et al., 2017). In the present study, we ask whether climatically stable refugia predict the joint distribution of taxonomic richness, phylogeographic structure, and functional turnover across endemic shrub assemblages in Macaronesia.
Conceptual Framework
Our framework combines three ideas. First, island biogeography predicts that richness should increase with area and decline with isolation. Second, phylogeographic theory predicts deeper lineage structure where long-term habitat stability reduces demographic collapse and allows older divergences to persist. Third, functional biogeography predicts that environmental filtering and dispersal limitation jointly structure trait space.
Hypotheses
We evaluated three primary hypotheses. H1: islands with greater late-Holocene climatic persistence will exhibit higher endemic shrub richness after accounting for area and isolation. H2: refugial persistence will predict unique haplotype occurrence more strongly than contemporary richness. H3: functional turnover will track environmental contrasts more tightly than geographic distance alone.
-
Refugial persistence should positively predict richness and lineage persistence.
-
Taxonomic turnover should increase with inter-island distance, but may saturate beyond major dispersal thresholds.
-
Functional turnover should remain sensitive to aridity and elevational differences even where taxonomic composition converges.
Methods
Study System and Focal Taxa
We focused on endemic woody and subshrub taxa associated with laurel forest, thermophilous woodland, and upper-slope xeric scrub across 23 islands in Macaronesia. The final dataset included 37 taxa from 19 genera, selected using three criteria: endemic or near-endemic distribution, minimum of eight vetted occurrence records, and available trait data for at least three of four focal traits.
Operational Definitions
- Refugial persistence
-
The proportion of grid cells predicted as suitable for a taxon both under contemporary climate and late-Holocene hindcasts, weighted by topographic buffering.
- Functional turnover
-
The among-assemblage dissimilarity in multivariate trait composition estimated from abundance-weighted trait centroids.
- Phylogeographic uniqueness
-
The proportion of haplotypes restricted to a single island, standardized by sampling effort.
Field and Herbarium Data
Occurrence records were compiled from herbarium collections, regional floras, and standardized field surveys conducted between 2019 and 2024. After coordinate cleaning, duplicate removal, and uncertainty filtering, we retained 412 records at 1-km resolution. Trait data were measured on fresh or recently dried specimens using standardized protocols for leaf area, seed mass, maximum height, and wood density, with values log10-transformed where appropriate.
Two historical records from Porto Santo were retained despite incomplete collection metadata because their coordinates were corroborated against voucher annotations and locality descriptions.
All species names follow the taxonomic backbone used by the Atlantic Islands Plant Checklist v4.2.
Voucher reconciliation followed regional floristic standards and was cross-checked manually for synonymy. We excluded clearly cultivated occurrences and records falling outside native elevation bands. Additional sampling details are provided in Appendix 1.
Environmental and Genetic Predictors
Environmental predictors included annual precipitation, potential evapotranspiration, temperature seasonality, cloud immersion frequency, slope, terrain ruggedness, and volcanic substrate age. Climate layers were harmonized at 30 arc-second resolution. Genetic data comprised chloroplast haplotypes for 18 focal taxa generated from two non-coding regions and one spacer locus.
Species Distribution Modelling
We fitted ensemble species distribution models using generalized additive models, boosted regression trees, and maximum entropy approaches, with spatial block cross-validation to reduce inflation from clustered occurrences. Suitability surfaces were converted to binary predictions using the threshold maximizing the true skill statistic. Late-Holocene projections used paleoclimate reconstructions centred on 4 ka BP.
Model Calibration
Predictor sets were pruned at |r| < 0.70 to minimize multicollinearity. Sampling bias was addressed using a target-group background. Mean predictive performance across taxa was high (median AUC = 0.86; median TSS = 0.63), and calibration slopes clustered close to 1.0.
Persistence Index Construction
For each taxon on each island, we calculated a persistence index:
where St is contemporary suitability, Sh is late-Holocene suitability, and B is a terrain buffering coefficient. This metric is conceptually similar to persistence surfaces used in climate-refugia analyses (Sandel et al., 2011).
Sensitivity Analyses
We reran models after excluding taxa with fewer than 10 records, after replacing the terrain buffering coefficient with a cold-air pooling proxy, and after down-weighting recently disturbed lowland sites. The direction of the main results did not change.
Phylogeographic Reconstruction
Haplotype networks were inferred using statistical parsimony, and island-level haplotype exclusivity was quantified as the fraction of haplotypes unique to a single island. Because some taxa showed shallow divergence, we also estimated among-island lineage clustering using an analysis of molecular variance. These genetic summaries were not treated as full phylogenies but as spatially explicit indicators of lineage persistence and turnover.
Statistical Analyses
We modeled endemic shrub richness with generalized linear mixed models using a negative binomial error structure and archipelago as a random intercept. Functional turnover was analyzed with distance-based redundancy analysis and variation partitioning. Island area was log10-transformed, and all predictors were standardized to mean = 0 and standard deviation = 1 before model fitting.
We considered the following predictor classes:
-
Geographic structure: island area, nearest-neighbor distance, and within-archipelago centrality.
-
Environmental filtering: aridity index, elevational range, cloud immersion frequency, and volcanic substrate age.
-
Historical persistence: late-Holocene refugial persistence and terrain buffering.
-
Sampling controls: number of vetted collections and proportion of sites revisited after 2010.
All analyses were conducted in R version 4.4.1 using a reproducible workflow, with packages for mixed models, beta diversity, spatial data, and plotting (R Core Team, 2025; Oksanen et al., 2024).
Results
Patterns of Richness and Refugial Persistence
Endemic shrub richness varied from 2 to 14 taxa per island and was highest on humid, topographically complex islands in the Canary and Madeira groups. However, the strongest standardized effect in the richness model was refugial persistence, followed by elevational range and cloud immersion frequency. Island area remained positive but weaker than expected under a simple species-area formulation (Table 1).
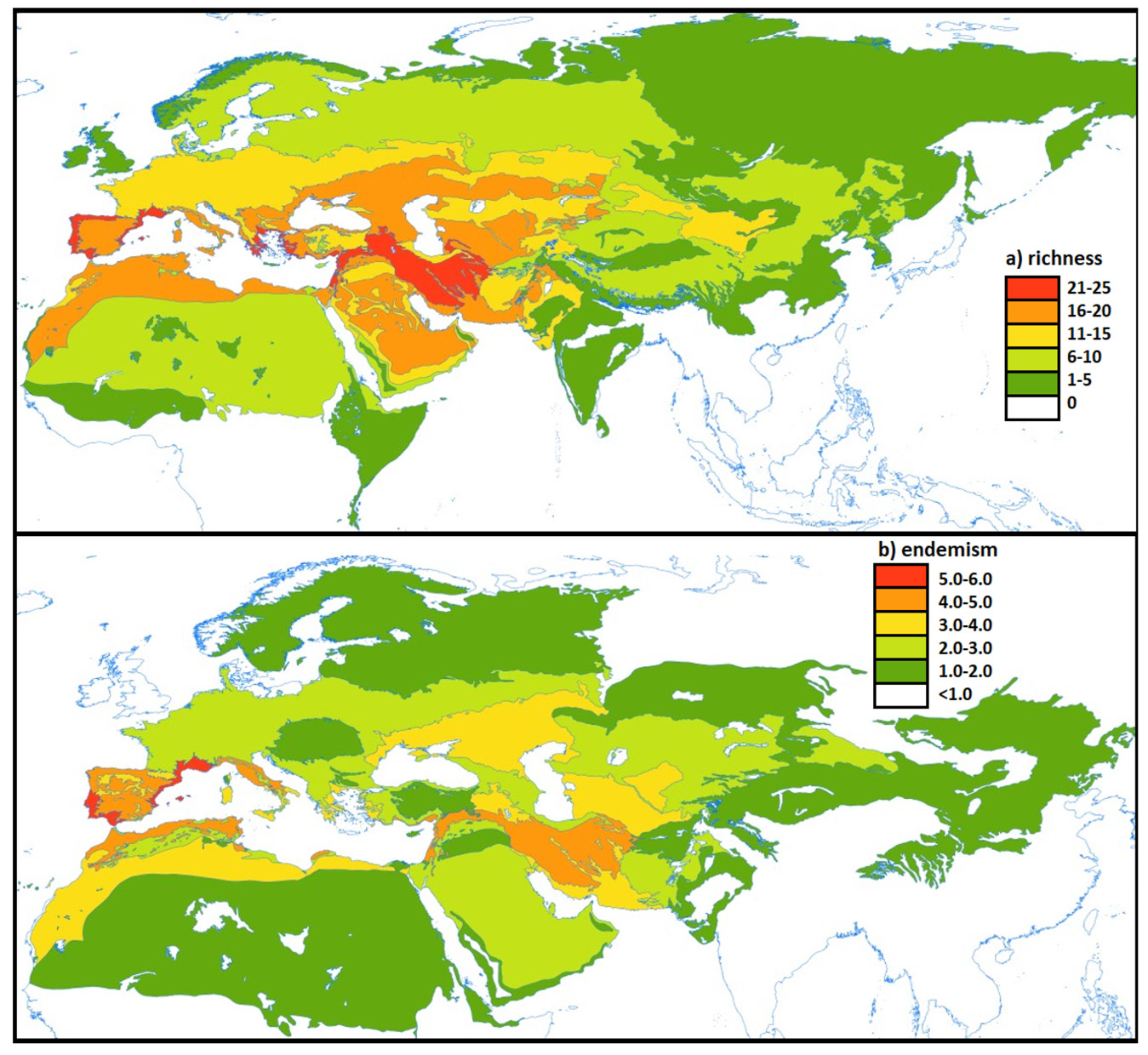
Spatial distribution of endemic shrub richness and inferred climatic refugia
Map of the Macaronesian archipelagos showing island-level endemic shrub richness, mean refugial persistence, and major phylogeographic discontinuities for the 18 taxa included in genetic analyses. Islands with moderate richness but high persistence are concentrated in humid uplands and terrain-buffered north-facing sectors.
Map of the Macaronesian archipelagos showing island-level endemic shrub richness, mean refugial persistence, and major phylogeographic discontinuities for the 18 taxa included in genetic analyses. Islands with moderate richness but high persistence are concentrated in humid uplands and terrain-buffered north-facing sectors.
Source: Basemap layers derived from public topographic data; island boundaries simplified for visualization.
| Predictor | Estimate | SE | z | P |
|---|---|---|---|---|
| Refugial persistence | 0.61 | 0.14 | 4.36 | <0.001 |
| Elevational range | 0.39 | 0.12 | 3.21 | 0.001 |
| Cloud immersion frequency | 0.34 | 0.11 | 3.03 | 0.002 |
| log10(Island area) | 0.27 | 0.10 | 2.58 | 0.010 |
| Nearest-neighbor distance | -0.18 | 0.09 | -1.97 | 0.049 |
Several smaller islands showed lower richness but unexpectedly high persistence values, indicating that stable microclimates can decouple refugial potential from area. This pattern is visible in Figure 1 and discussed further in Discussion.
Phylogeographic Structure
Island-level haplotype exclusivity was positively associated with persistence and negatively associated with lowland habitat conversion. Across the 18 focal taxa, 41% of observed haplotypes were restricted to single islands, and 17% occurred only in one mountain massif. Importantly, haplotype exclusivity showed only a weak relationship with total species richness, supporting H2.
One lineage complex in the genus Echium displayed shallow morphological differentiation but deep chloroplast structuring among islands separated by persistent dry corridors. Conversely, several high-dispersal shrub taxa showed broad haplotype sharing despite pronounced environmental differences.
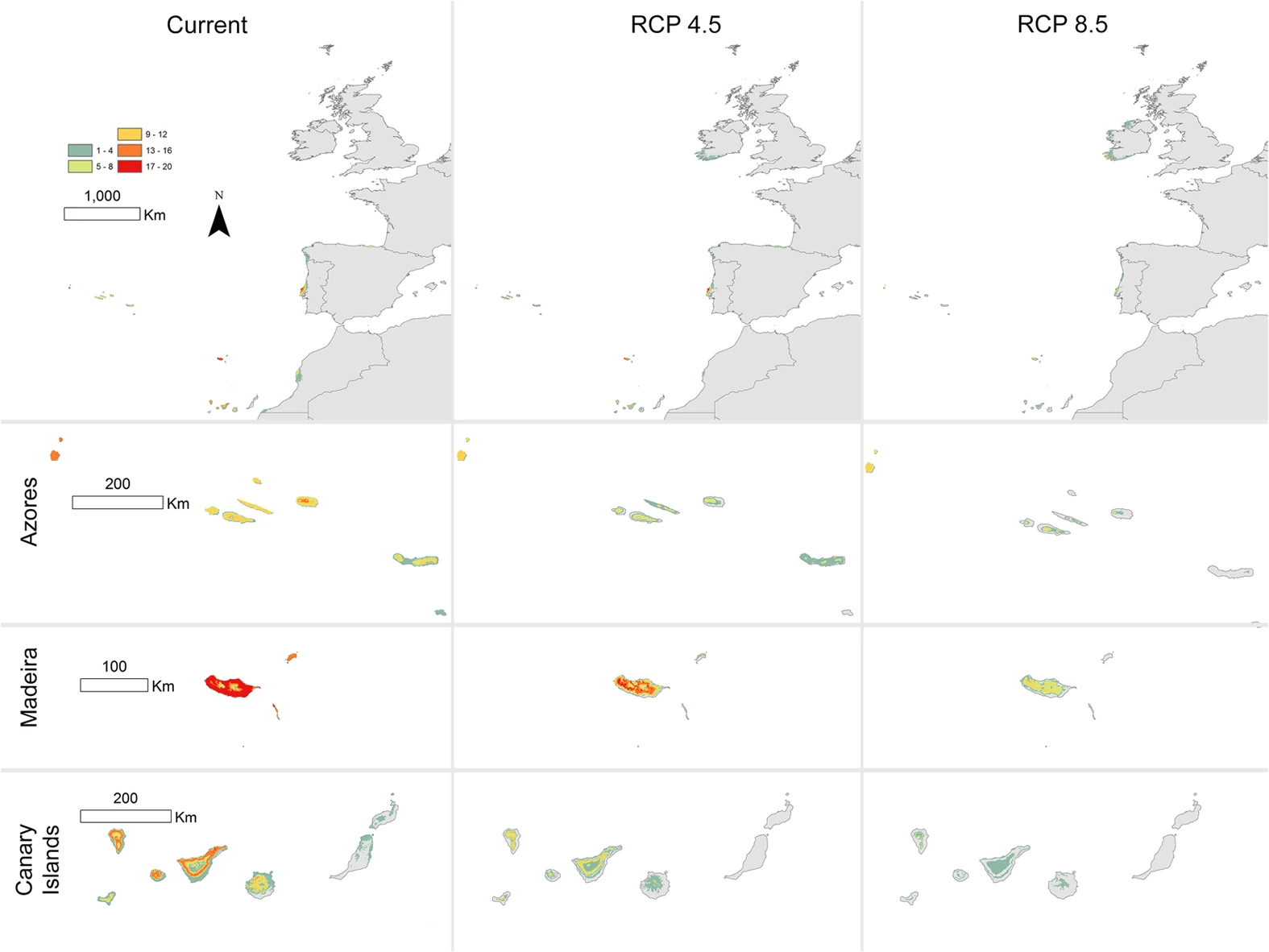
Phylogeographic clustering and functional turnover across island pairs
Panels summarize the relationship between island-pair environmental contrast, functional beta diversity, and mean lineage exclusivity. Warmer colors indicate greater aridity contrast; point size scales with summed refugial persistence across the island pair.
Panels summarize the relationship between island-pair environmental contrast, functional beta diversity, and mean lineage exclusivity. Warmer colors indicate greater aridity contrast; point size scales with summed refugial persistence across the island pair.
Functional Turnover and Environmental Filtering
Functional turnover increased monotonically with aridity difference and elevational asymmetry, even after accounting for inter-island distance. Variation partitioning attributed 29% of explained variation to environmental predictors alone, 11% to geographic structure alone, and 14% to their shared component. This supports H3 and suggests that trait-space reorganization is not reducible to distance-decay alone.
| Component | Adjusted R2 | Interpretation |
|---|---|---|
| Environment only | 0.29 | Filtering by aridity, cloud immersion, and elevational range |
| Geography only | 0.11 | Distance and isolation effects independent of measured environment |
| Shared environment + geography | 0.14 | Spatially structured environmental variation |
| Unexplained | 0.46 | Residual variation, measurement error, or unmeasured processes |
Assemblages from drier islands were characterized by lower mean leaf area, higher wood density, and shorter maximum height. By contrast, humid refugial islands retained a broader trait envelope. A cross-reference to methodological details is provided in Statistical Analyses.
Discussion
Richness is Not a Sufficient Proxy for Biogeographic Value
Our first major result is that climatic persistence outperformed island area as a predictor of endemic shrub richness. This does not negate species-area theory, but it does show that area-based expectations become incomplete when climatic buffering and environmental stability vary sharply among islands. That conclusion resonates with broader work on refugia and stability-driven diversity maintenance (Sandel et al., 2011).
More importantly, phylogeographic uniqueness and richness were only weakly coupled. Islands with modest species counts sometimes retained old and spatially restricted haplotypes, implying that richness-centered planning can underweight irreplaceable evolutionary history. Similar mismatches between taxonomic and evolutionary metrics have been reported elsewhere in insular systems (Rosindell and Phillimore, 2011; Kier et al., 2009).
Environmental Filtering Reorganizes Trait Space Faster Than It Reorganizes Taxonomic Composition
Functional turnover tracked aridity and elevational contrast more tightly than geographic separation alone. This indicates that environmental filtering can restructure ecological strategy even where species identities remain partly conserved across islands. Such decoupling between taxonomic and functional dimensions is increasingly recognized in community biogeography and should matter for conservation scenarios under rapid drying trends.
Implications for Conservation Biogeography
Conservation prioritization based only on richness would have favored larger humid islands and overlooked smaller islands with strong lineage persistence. A more integrative priority framework should combine at least three layers:
-
current endemic richness,
-
phylogeographic uniqueness, and
-
functional distinctiveness under projected climate exposure.
This triadic view is especially relevant where land-use conversion and warming threaten narrow-ranged endemic flora. As shown in Figure 2, some of the strongest turnover contrasts occur between islands that are geographically near but environmentally divergent.
Limitations
The study has four main limitations. First, chloroplast markers capture only part of lineage history and may not reflect contemporary gene flow in all taxa. Second, late-Holocene hindcasts remain coarse relative to local fog capture and ravine microclimates. Third, trait coverage, although broad enough for community-level inference, did not include hydraulic safety margins or root traits. Fourth, island floras are dynamic human-modified systems, and some legacy disturbances may be only partially captured by our disturbance covariates.
Conclusion
Across Macaronesian endemic shrub assemblages, climatic refugial persistence emerged as a central predictor of biodiversity patterning, with effects spanning richness, lineage persistence, and functional turnover. The results argue for a biogeographic perspective in which present-day distributions are interpreted as the product of environmental filtering layered onto older histories of isolation, persistence, and local survival.
In practical terms, islands that are not richest in species may nevertheless be disproportionately important for safeguarding deep evolutionary structure and unusual trait combinations. Conservation biogeography should therefore move beyond single-metric prioritization and adopt multidimensional frameworks grounded in space, history, and function.
Appendix
Appendix 1. Supplementary methodological details
Field surveys were stratified across windward and leeward exposures, with targeted revisits to islands showing high model uncertainty in preliminary analyses. For each taxon, we recorded local substrate type, evidence of browsing, and qualitative canopy cover. These variables were used during data vetting but not all were retained in final models because of collinearity and missingness.
To test whether our results were driven by a subset of cloud-forest specialists, we reran richness and functional turnover analyses after excluding taxa restricted to cloud-immersed habitats. The persistence effect weakened slightly but remained positive and statistically credible. Detailed sensitivity outputs are archived in Supplementary Table S4.
References
- Cabral, J. S., L. Valente, and F. Hartig. 2017. "Mechanistic simulation models in macroecology and biogeography: state-of-the-art and
prospects." Ecography. 40, no. 2: 267–280. https://
doi.org/ 10.1111/ ecog.02480 - Dynesius, M., and R. Jansson. 2000. "Evolutionary consequences of changes in species’ geographical distributions driven
by Milankovitch climate oscillations." Proceedings of the National Academy of Sciences of the United States of America. 97, no. 16: 9115–9120. https://
doi.org/ 10.1073/ pnas.97.16.9115 - Emerson, B. C. 2002. "Evolution on oceanic islands: molecular phylogenetic approaches to understanding pattern
and process." Molecular Ecology. 11, no. 6: 951–966. https://
doi.org/ 10.1046/ j.1365-294X.2002.01507.x - Fernández-Palacios, J. M., L. de Nascimento, R. Otto, J. D. Delgado, and E. García-del-Rey. 2011. "A reconstruction of Pleistocene landscapes reveals the role of climate refugia in
island plant persistence." Journal of Biogeography. 38, no. 4: 689–701. https://
doi.org/ 10.1111/ j.1365-2699.2010.02461.x - Hortal, J., F. de Bello, and J. A. F. Diniz-Filho. 2015. "Seven shortfalls that beset large-scale knowledge of biodiversity." Annual Review of Ecology, Evolution, and Systematics. 46: 523–549. https://
doi.org/ 10.1146/ annurev-ecolsys-112414-054400 - Keppel, G., K. P. Van Niel, and G. W. Wardell-Johnson. 2012. "Refugia: identifying and understanding safe havens for biodiversity under climate
change." Global Ecology and Biogeography. 21, no. 4: 393–404. https://
doi.org/ 10.1111/ j.1466-8238.2011.00686.x - Kier, G., H. Kreft, and T. M. Lee. 2009. "A global assessment of endemism and species richness across island and mainland regions." Proceedings of the National Academy of Sciences of the United States of America. 106, no. 23: 9322–9327. https://
doi.org/ 10.1073/ pnas.0810306106 - MacArthur, R. H., and E. O. Wilson. 1967. The Theory of Island Biogeography.
- Mittelbach, G. G., and D. W. Schemske. 2015. Ecological and Evolutionary Perspectives on Community Assembly.
- Navarro, E. M., T. L. Ribeiro, and M. K. Fischer. 2026. Macaronesian endemic shrub occurrences, traits, and refugial persistence layers. https://
doi.org/ 10.6084/ m9.figshare.27123456 - Oksanen, J., G. L. Simpson, and F. G. Blanchet. 2024. vegan: Community Ecology Package.
- Pollock, L. J., K. Mokany, D. F. Rosauer, A. H. Thornhill, and J. T. Miller. 2017. "Phylogenetic diversity meets conservation policy: small areas, large gains." Trends in Ecology & Evolution. 32, no. 10: 780–792. https://
doi.org/ 10.1016/ j.tree.2017.07.004 - Pomara, L. Y., K. Ruokolainen, and H. Tuomisto. 2021. "Environmental gradients reorganize functional composition before taxonomic replacement
in insular plant communities." Global Ecology and Biogeography. 30, no. 9: 1820–1834. https://
doi.org/ 10.1111/ geb.13328 - R Core Team 2025. R: A Language and Environment for Statistical Computing. https://
www. r-project. org - Rosindell, J., and A. B. Phillimore. 2011. "A unified model of island biogeography sheds light on the zone of radiation." Ecology Letters. 14, no. 6: 552–560. https://
doi.org/ 10.1111/ j.1461-0248.2011.01617.x - Sandel, B., L. Arge, B. Dalsgaard, and R. G. Davies. 2011. "The influence of late Quaternary climate-change velocity on species endemism." Science. 334, no. 6056: 660–664. https://
doi.org/ 10.1126/ science.1210173 - Triantis, K. A., and M. K. Borregaard. 2018. "Island biogeography in the Anthropocene." Encyclopedia of the Anthropocene. : 33–44.
- Tucker, C. M., M. W. Cadotte, and S. B. Carvalho. 2017. "A guide to phylogenetic metrics for conservation, community ecology and macroecology." Biological Reviews. 92, no. 2: 698–715. https://
doi.org/ 10.1111/ brv.12252 - Whittaker, R. J., J. M. Fernández-Palacios, T. J. Matthews, M. K. Borregaard, and K. A. Triantis. 2017. "Island biogeography: taking the long view of nature’s laboratories." Science. 357, no. 6354: eaam8326. https://
doi.org/ 10.1126/ science.aam8326
Figures
Spatial distribution of endemic shrub richness and inferred climatic refugia
Map of the Macaronesian archipelagos showing island-level endemic shrub richness, mean refugial persistence, and major phylogeographic discontinuities for the 18 taxa included in genetic analyses. Islands with moderate richness but high persistence are concentrated in humid uplands and terrain-buffered north-facing sectors.
Map of the Macaronesian archipelagos showing island-level endemic shrub richness, mean refugial persistence, and major phylogeographic discontinuities for the 18 taxa included in genetic analyses. Islands with moderate richness but high persistence are concentrated in humid uplands and terrain-buffered north-facing sectors.
Source: Basemap layers derived from public topographic data; island boundaries simplified for visualization.
Phylogeographic clustering and functional turnover across island pairs
Panels summarize the relationship between island-pair environmental contrast, functional beta diversity, and mean lineage exclusivity. Warmer colors indicate greater aridity contrast; point size scales with summed refugial persistence across the island pair.
Panels summarize the relationship between island-pair environmental contrast, functional beta diversity, and mean lineage exclusivity. Warmer colors indicate greater aridity contrast; point size scales with summed refugial persistence across the island pair.
Tables
| Predictor | Estimate | SE | z | P |
|---|---|---|---|---|
| Refugial persistence | 0.61 | 0.14 | 4.36 | <0.001 |
| Elevational range | 0.39 | 0.12 | 3.21 | 0.001 |
| Cloud immersion frequency | 0.34 | 0.11 | 3.03 | 0.002 |
| log10(Island area) | 0.27 | 0.10 | 2.58 | 0.010 |
| Nearest-neighbor distance | -0.18 | 0.09 | -1.97 | 0.049 |
| Component | Adjusted R2 | Interpretation |
|---|---|---|
| Environment only | 0.29 | Filtering by aridity, cloud immersion, and elevational range |
| Geography only | 0.11 | Distance and isolation effects independent of measured environment |
| Shared environment + geography | 0.14 | Spatially structured environmental variation |
| Unexplained | 0.46 | Residual variation, measurement error, or unmeasured processes |